Terror 25 Mate for Love or Status

https://www.google.com/search?q=dore+illustrations+for+raven&source=lnms&tbm=isch&sa=X&ved=0ahUKEwjbz_apjZTaAhUPM6wKHZSwBaoQ_AUICigB&biw=1260&bih=641#imgrc=9yXhz_7MjO5lyM: downloaded 3/30/18
It is the 15th of April 2018. The theory of the mechanism that links kinship and fertility, which is to say that program for which you have the source code, can give a number of patterns for the time course of population size over many generations.
The first is a constant population. That is sort of the skeleton in my closet. I have not been able to produce such a behavior although I have not pursued it with much vim nor have I pursued it after tweaking the program to limit the effect of pre-zygotic infertility. I am discouraged from the attempt because of the very noisy quality of my results. Using the computer at which I now sit, I could in theory increase the number of sites affecting infertility from 100 per segment to about 2,000 per segment in the arrays. That would reduce the noise, I expect, by a factor of 40 or 50, which probably would be enough. I entreat you to do it yourself.
A pattern I do find is damped oscillation:
Graphing offspring on the vertical axis against successive generations on the horizontal
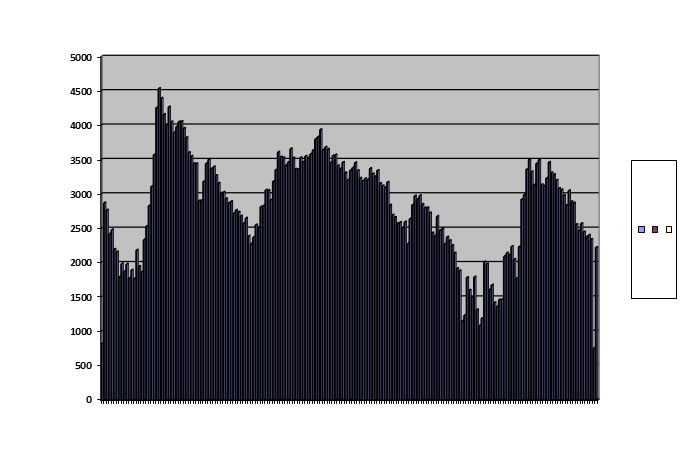
This curve was obtained with a run in which only 200 of the saved population were brought out and run; all other conditions were identical with the baseline. This is an ungainly example, but it can be seen in wild voles, as we have seen, and in captive fruit flies. As lest so far as our data goes, these animals never exhibit any other pattern although they have the opportunity. This pattern is a winner. I have never seen it in a population that went extinct.
A second pattern is the double peak:
Conditions were identical with those of the baseline run except that maximum population was permitted to rise to 3,000.
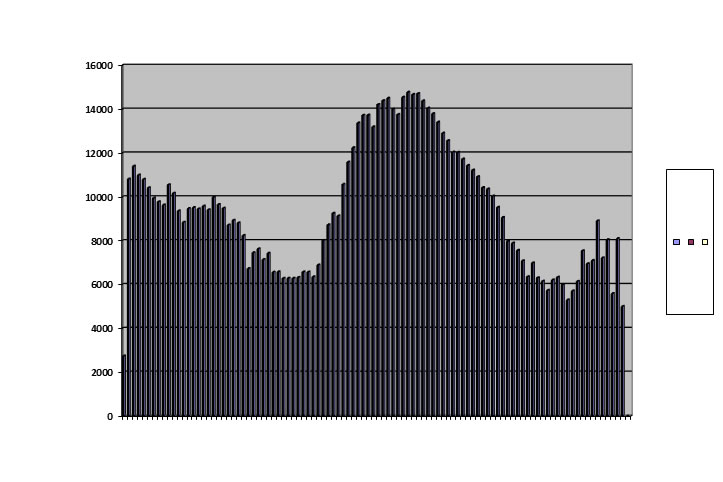
We have seen this double peak in the survival of civilizations and dynasties, in the farmers of Long House Valley, and in wild mouse populations in Australia. None of these, neither human nor murine, showed damped oscillation in spite of ample opportunity. This pattern is a loser. Extinction always supervenes.
A final pattern is a single peak and a crash:
I let the maximum population rise to 4,000, but otherwise kept conditions identical with the baseline run and the run with two peaks.
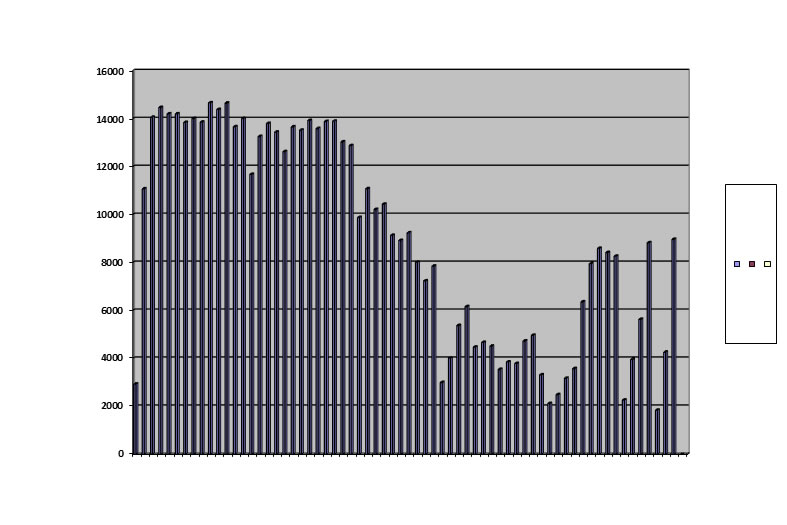
It died out after 67 generations. Ignoring the abortive final rally, the pattern looks like fast up, slow down, tends to level off and then collapse. This pattern has been seen in Canadian weevils, captive mice and wild mice. It is a loser, always leading to extinction.
So we can clump animals into two kinds, those that have damped oscillation and those that have the double peak. Allow me to speculate on the difference.
There are two ways animals are attracted to each other: they may be attracted by kinship, which has been shown by Bateson in his Japanese quail, and they can be attracted by status, which has been shown by Calhoun and his mice. Forgive me if I phrase that in some animals mate selection is based predominately on love and in others it is based predominately on status. Quail and fruit flies look for love, humans and mice look for status.
This suggestion is frivolous, but not entirely frivolous. In theory it could be tested. Imagine some arrangement whereby every fly in a fruit fly colony could be traced throughout it’s life for a few generations. Possibly an MRI study run twenty-four, seven could be done without cooking the flies. The result would have to be analyzed by computer, and the cost would be prodigious. But in the event such a study were practicable, the prediction would be that the flies would have a relatively high frequency of forming mating pairs with nearer cousins. In fact I shall risk ridicule and say that they would form these bonds while they were still maggots.
YouTube video scripts directory
Home page